Understanding flower development involves recognizing diverse floral structures and the genetic models that explain their formation. Flowers can be classified based on the presence of their parts and their reproductive structures.
A complete flower is one that possesses all four main whorls of floral organs: sepals, petals, stamens, and carpels. The image below illustrates the typical anatomy of such a flower.
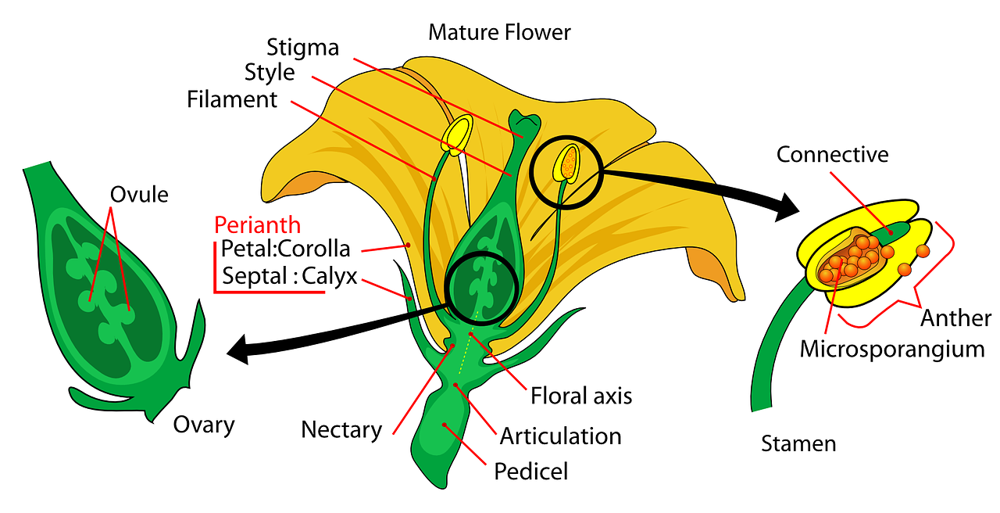
Figure 1: General anatomy of a complete flower, showing key reproductive and accessory organs.
The specification of these floral organs is largely governed by the ABCE model. In the classic ABCE model, distinct classes of homeotic genes interact combinatorially to determine the identity of the four floral organ types arranged in concentric whorls:
- Sepals: A-function genes alone (plus E-function).
- Petals: A-function and B-function genes together (plus E-function).
- Stamens: B-function and C-function genes together (plus E-function).
- Carpels: C-function genes alone (plus E-function).
The E-function genes (often SEPALLATA genes) are generally required for the proper development of all four whorls. The A-function and C-function genes are typically mutually antagonistic.
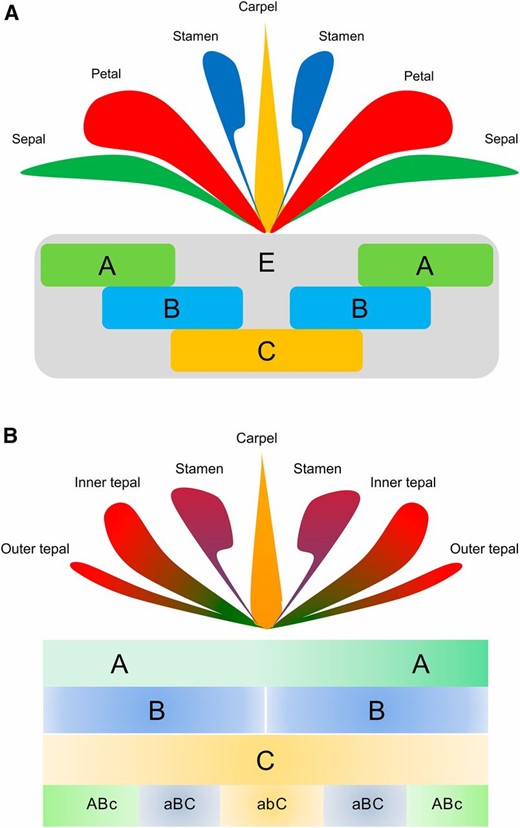
Figure 2: Comparison of the classic ABCE model (A) and the fading borders model (B) of floral organ identity. Modified from Chanderbali et al. (2016).[3]
While the classic ABCE model (Figure 2A) defines sharp boundaries, the "fading borders model" (Figure 2B)[3] suggests more gradual transitions in gene activity, potentially leading to morphologically intergrading organs like tepals, where gene class influence might be lower (indicated by lowercase letters like "aBC").
An incomplete flower is one that lacks one or more of the four main floral whorls. A prime example is found in the Poaceae family (grasses), such as rice. These flowers are highly specialized for wind pollination.
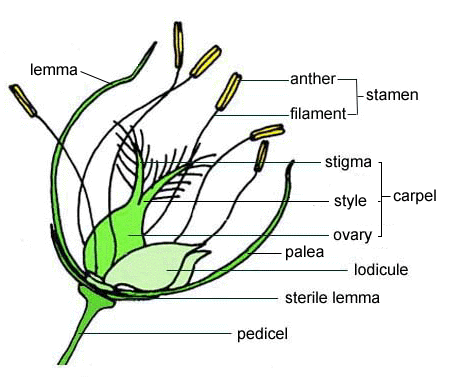
Figure 3: Structure of a rice flower (floret of Poaceae), an example of an incomplete flower adapted for wind pollination.
Key features of a grass floret include:
- Reduced Perianth: The sepals and petals are typically absent or highly reduced to small, scale-like structures called lodicules, which help in opening the flower.
- Protective Bracts: The reproductive organs are enclosed by specialized bracts: the lemma and the palea. Additional sterile lemmas or glumes may be present at the base.
- Wind Pollination Adaptations: Stamens often have long, pendulous filaments and large anthers to release copious amounts of pollen into the wind. Stigmas are typically large, feathery, and branched to effectively capture airborne pollen.
Unisexual flowers (also called imperfect flowers) possess either stamens (male reproductive parts) or carpels/pistils (female reproductive parts), but not both. Pumpkin (Cucurbita pepo) is a classic example of a plant with unisexual flowers, typically being monoecious (having separate male and female flowers on the same plant).
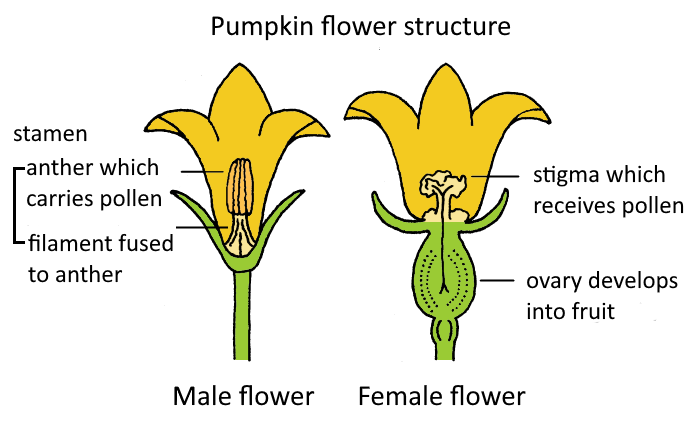
Figure 4: Structure of pumpkin (Cucurbita pepo) male and female flowers, illustrating unisexual flower characteristics.
Distinguishing features:
- Male Flower: Contains only stamens, which produce pollen. The filaments of stamens in pumpkin are often fused. After releasing pollen, the male flower typically withers.
- Female Flower: Contains only a pistil (composed of stigma, style, and ovary). A key identifying feature is the presence of a prominent inferior ovary at the base of the flower, which looks like a miniature pumpkin. This ovary, if pollinated, will develop into the fruit.
Pollination in pumpkin is usually carried out by insects, such as bees, which transfer pollen from male flowers to female flowers.
Plants are broadly classified into monocotyledons (monocots) and dicotyledons (dicots or eudicots). Their flower structures often exhibit distinct characteristics. The image below shows a typical dicot flower structure.
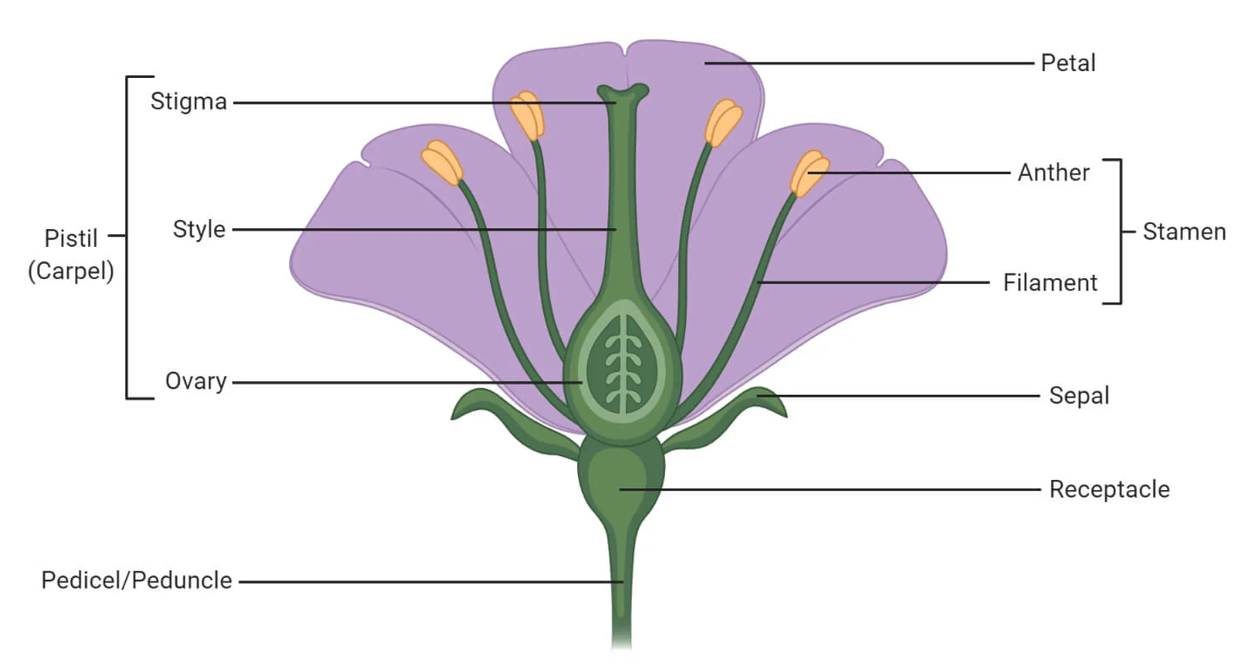
Figure 5: General structure of a dicotyledon flower.
Typical characteristics of dicot flowers include:
- Floral Parts in 4s or 5s: The number of sepals, petals, and stamens is often in multiples of four or five.
- Differentiated Perianth: There is usually a clear distinction between sepals (forming the calyx) and petals (forming the corolla).
- Net-like Leaf Venation: While not a floral trait, dicots typically have leaves with reticulate (net-like) venation.
- Taproot System: Dicots often possess a primary taproot.
In contrast, monocot flowers (like grasses or lilies) typically have floral parts in multiples of three, often undifferentiated perianth (tepals), parallel leaf venation, and a fibrous root system.
Epigenetic mechanisms, as discussed below, play a crucial role in establishing and maintaining the precise expression domains of genes like those in the ABCE model, thereby ensuring correct floral patterning across these diverse flower types.